
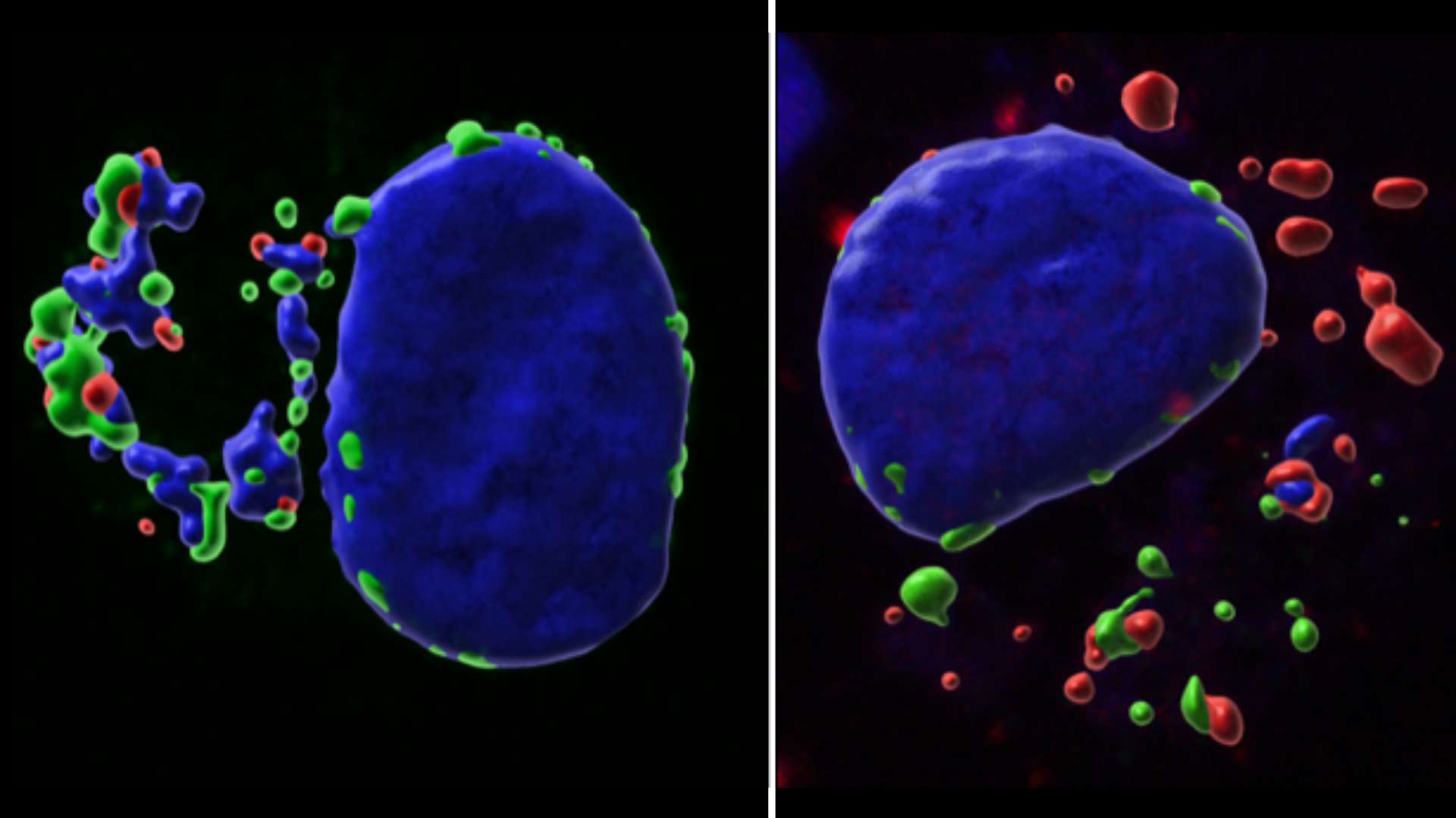
L’équipe de recherche du Pr Jonathan Weitzman a mis en lumière le mécanisme par lequel le parasite Theileria annulata, responsable de maladies semblables aux cancers chez les bovins, échappe au mécanisme de défense de la cellule hôte.
ERC Advanced Grant 2023 : le professeur Anton Zorich, mathématicien à l’université Paris Cité compte parmi les 37 lauréates et lauréats français.


Actualités














Offre de formation
La réussite de tous les étudiants, et ce dès leur entrée à l’université, est une priorité pour Université Paris Cité.
Agenda

29 avril 2024

UCLouvain et en ligne
30 avril 2024

Campus des Grands Moulins, Amphi Buffon
17 mai 2024
En visioconférence

22 mai 2024

campus Saint-Germain-des-Prés


14 juin 2024
En visioconférence
